
![]()
2020年10月13日
国立研究開発法人 国立精神・神経医療研究センター (NCNP)
国立大学法人 京都大学
印刷用PDF(742KB)
腕の自由自在な動きをつくりだす多機能な神経細胞群の発見
~運動の方向と大きさを同時にコントロールする神経メカニズムの解明~

国立研究開発法人 国立精神・神経医療研究センター(NCNP)神経研究所、モデル動物開発研究部のAmit Yaron研究員、David Kowalski訪問研究員と関和彦部長、京都大学白眉センター・医学研究科の武井智彦特定准教授は、自由自在な動きが特徴の霊長類の手や腕が、脊髄にある多機能な運動細胞群によってコントロールされていることを発見しました。
例えば私たちの手には、27種類の筋肉と18個の関節があります。私たちの神経系はこれらをうまく組み合わせて運動制御していますが、組み合わせパタンは極めて膨大でそのリアルタイムな制御は最新のスーパーコンピューターを用いても実現できません。神経系がどのようにこの計算を行なっているのかは未知ですが、その一つの解決法として「運動モジュール」仮説*1が広く認められています。これは、神経系は個々の筋肉や関節を直接操作するのでなく、脊髄*2にある少数の神経細胞群(運動モジュールと呼ばれる)を組み合わせて制御しているという仮説です。この仮説はこれまでカエルを中心にその証拠が示されてきましたが、ヒトを含めた霊長類においては証明されていませんでした。
そこで、本研究グループでは、上記のカエルを対象とした先行研究の実験手法を応用して、マカクサルの脊髄においてこの「運動モジュール」を同定する方法を開発しました。つまり、腕の運動に関わる脳からの信号を筋肉に伝える中継点である脊髄に多数の刺激電極を埋め込み、電気刺激とそれによって引き起こされた筋肉の活動の関連性の分析から、「運動モジュール」に相当する神経細胞群を発見する方法です。そして、この方法を用いて霊長類の脊髄に初めてその神経細胞群を発見しただけでなく、カエルと異なり霊長類の「運動モジュール」は運動の方向と大きさを別々に制御し、かつ同時にできることを発見しました。本研究は霊長類の手や腕の運動制御メカニズムを解明しただけでなく、その異常にともなう様々な運動障害の解明やリハビリへの応用、また効率的なロボットハンドの制御など様々な社会的波及効果があると考えられます。
この研究成果は、日本時間2020年10月13日(火)午前5時(米国東部標準時 2020年 10月 12日午後3時)に米国科学アカデミー紀要 (PNAS) オンライン版に掲載されました。
研究の背景
私たちの日常生活において、腕の働きは驚くほど多様です。私たちの神経系は右や左、また近くや遠くなど、様々な運動の方向と大きさの組み合わせによって、この多様な腕運動を作り出していると考えられます。しかし、このような多様な運動の方向と運動の大きさの制御が行われる神経メカニズムは長年脳科学における謎であり、いくつかの仮説が提唱されていますが、現在も解明されていません。
その仮説の一つが、運動モジュール仮説です。神経系には運動の方向(筋活動の組み合わせ)を表現する神経細胞群(モジュール)が少数存在し、脳はそれらをいろんな割合で組み合わせて使うことによって、多様な動きの方向や大きさを作り出しているという考えです。この考えを用いると、複雑なロボットの動きが少ない計算で実現でき、また多くの筋活動の組み合わせを簡単な数式で説明できることから、工学から臨床医学などで広く使われています。しかし、この運動モジュールはこれまで主としてカエルの神経系では実験的に発見されていましたが、実はヒトや霊長類の神経系では発見されていませんでした。
研究結果の概略
関部長らの研究グループはこの点に着目し、霊長類の神経系におけるこの運動モジュールの存在を実験的に検証しました。特に、歩行運動などの中枢がある脊髄に注目し、カエルで用いられていたものと同じ実験方法で実験を行いました。脊髄内の2つの異なった位置を同時に電気刺激*3して引き起こされる腕運動の方向は、それぞれを単独に刺激した際の運動方向の単純な線形和で説明可能なことを証明しました。この結果は、カエルの脊髄での実験結果と同一であり、サルの脊髄に運動方向を決めるモジュールが存在することが世界で初めて証明されました。次に、運動の大きさに注目すると、予想外の発見がありました。運動の方向と異なり、運動の大きさは複数の脊髄部位を同時に刺激すると単一刺激に比べて数倍から数十倍大きな運動が引き起こされたのです。この現象はカエルや齧歯類では報告されておらず、霊長類特有のものである可能性があります。そして、霊長類は、同じ運動モジュールを用いて、運動の方向と大きさを別々に制御していることを示唆する重要な発見でした。
研究の方法と結果
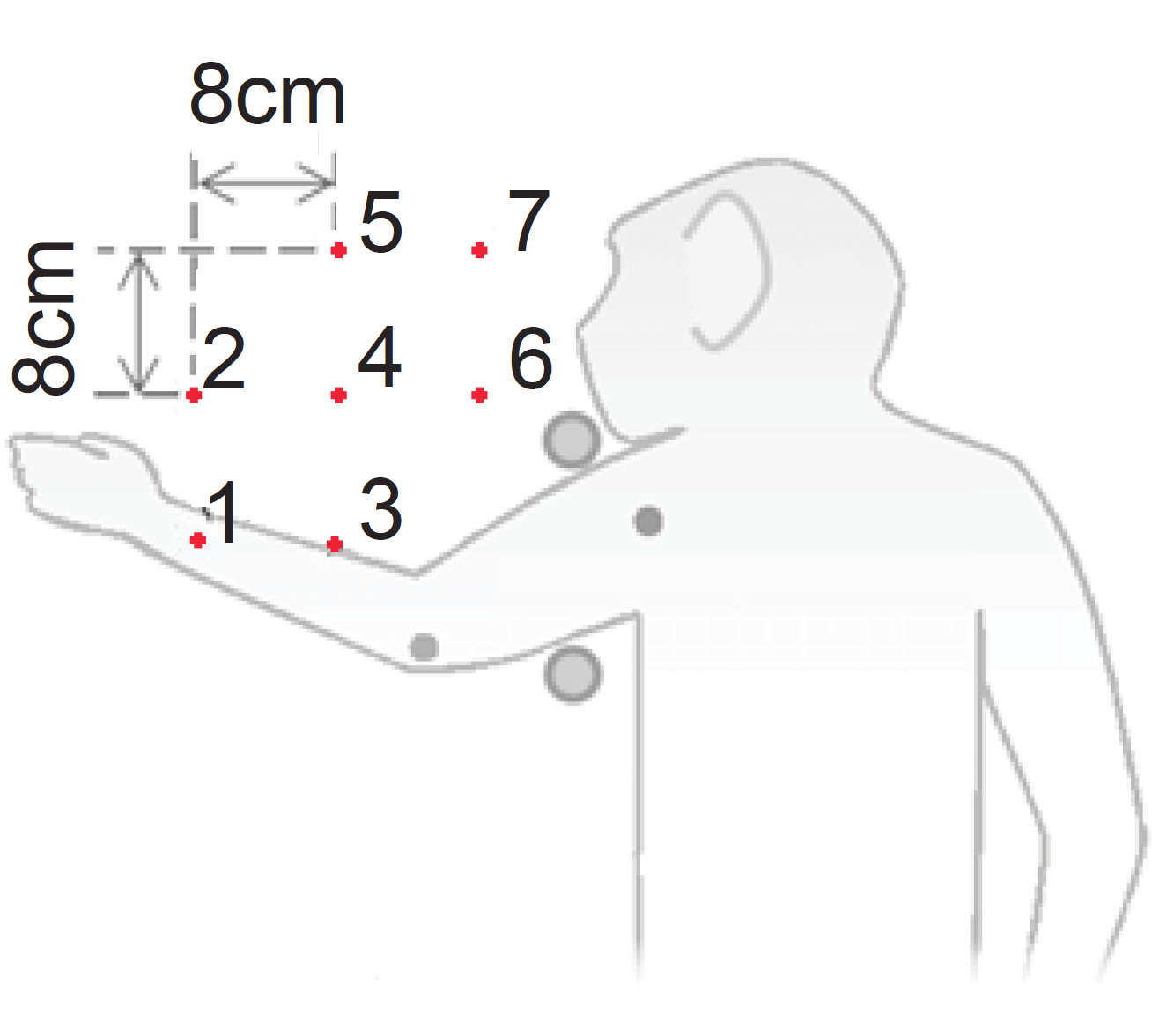
麻酔下のマカクサルの下位頸髄に16本の金属電極を慢性的に埋め込む手術を3頭の動物に対して行いました。手術から回復後、それぞれの動物の麻酔下で腕と身体を測定台に固定し(図2)、脊髄への電気刺激によって誘発される腕運動の方向と大きさを手首に力センサーを装着することによって計測しました(図3)。手首の位置を7点間で移動させると、脊髄刺激によって異なった方向の力ベクトルが計測されました(図4)。次に、二つの電極(電極AとB)への刺激によって発生する力ベクトルについて比較しました。Aを刺激すると図5A、またBを刺激すると図5Bのような方向へ力ベクトルが引き起こされました。次に、AとBを同時に刺激すると図5Dのような方向へ力ベクトルが引き起こされましたが、それはAとBのベクトルの線形和で想定される力発生パタン(図5C)とよく一致していました。このことから、霊長類の腕運動の方向は、脊髄における個々の運動モジュールの働きの単純な線形和*4で説明可能なことを示しました。同様な結果が3頭のサルの30電極ペアにおいて再現されたことから、霊長類の頸髄に運動モジュールが存在すると結論づけました。
図3:サルの上肢の力を手首部分で計測する力センサー
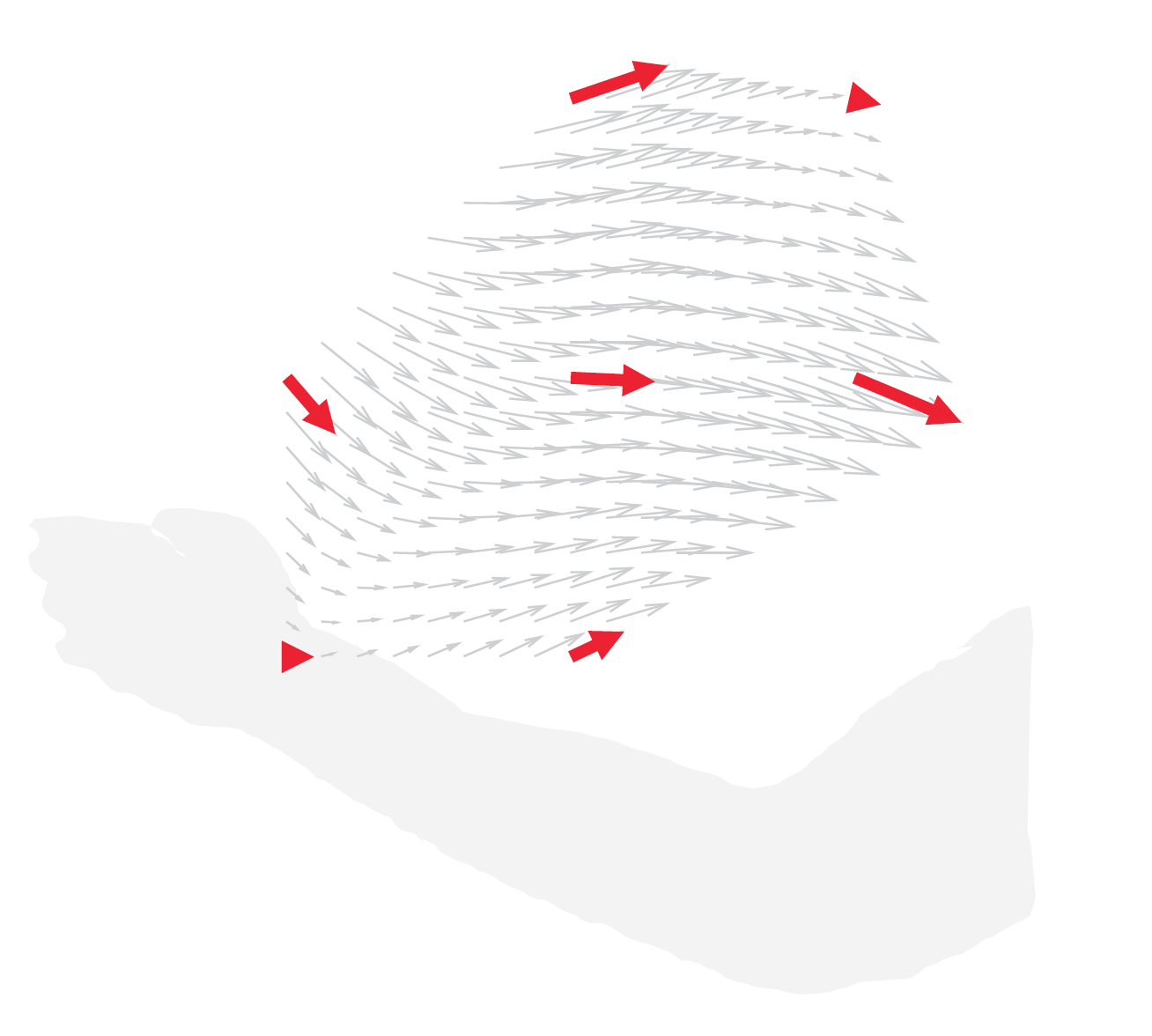
図4:脊髄刺激によって発生する力の方向は手首の位置によって異なった。
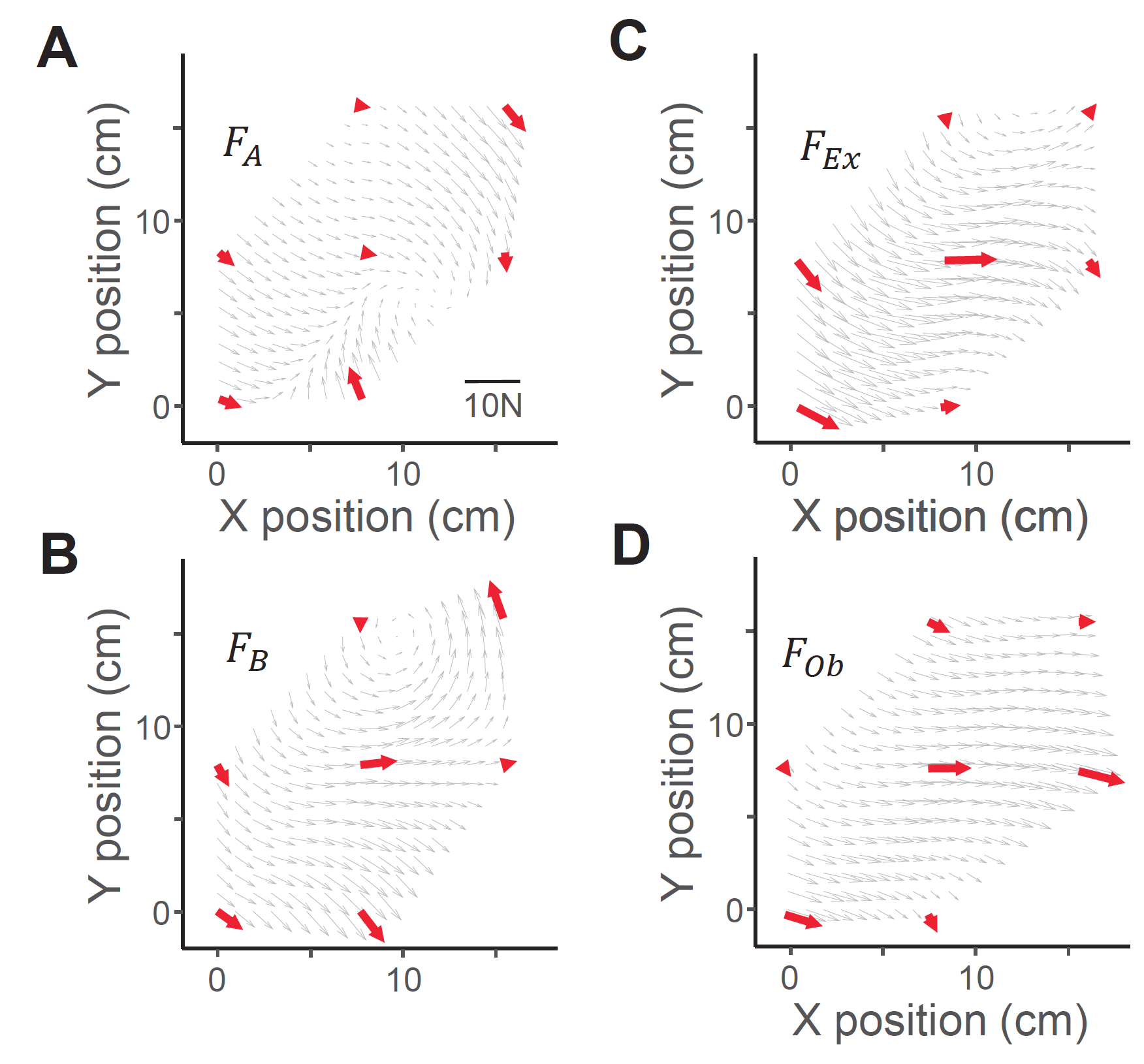
図5:A,Bはそれぞれ脊髄別部位を別々に刺激した例。別のパタンの力が観察される。C:AとBの線形和で想定される力パタン。D: AとBへの同時刺激によって観察された実際の力。CとDが近似していることから、脊髄部位AとBは「運動モジュール」と考えられる。
しかし、誘発された力の大きさについて同様な解析をすると、運動方向とは全く異なる結果が得られました。すなわち、脊髄2点刺激において誘発される運動は、個々の刺激で誘発される運動サイズの線形和*4(図6黄矢印)にはならず、線形和の数倍から数十倍大きな力が誘発されました(図6赤矢印:超線形和現象*4)。これは従来の報告にない、全く予想外の現象でした。では、霊長類の脊髄ではなぜ、四肢運動の大きさが運動モジュール出力の線形和で説明できないのか。本研究グループは、「この超線形和現象が霊長類に特有な手指の運動の多彩さと関係しているのではないか」、「そうだとすれば、手指の筋活動と肩肘の筋活動を別々に解析した場合、手指にこの超線形和現象が顕著に認められるのではないか」という新たな仮説を立てました。そして実際に筋電図を解析した結果、仮説通り手首や手指の筋において顕著に超線形和現象が現れることを見出しました(図7)。
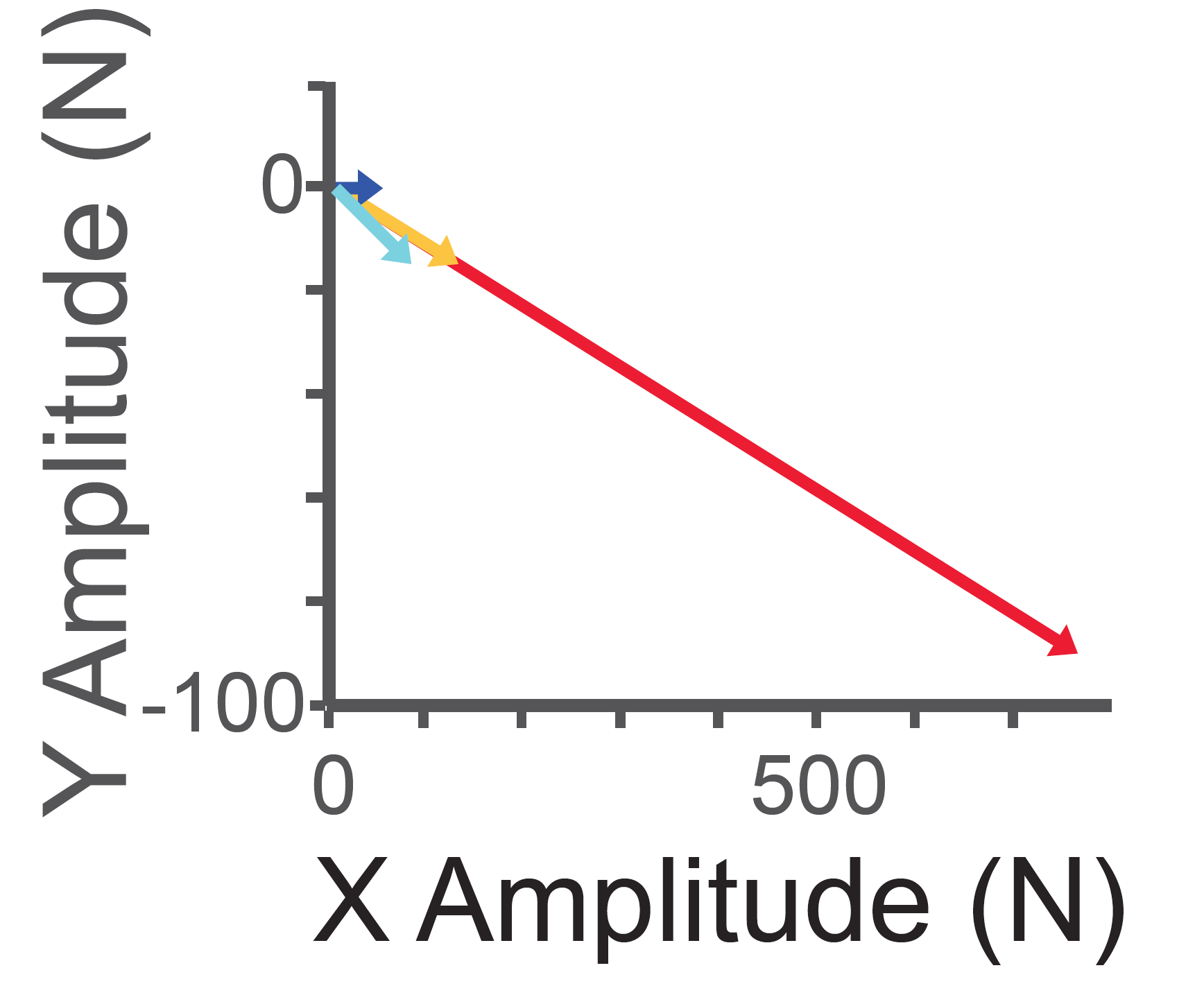
図6:異なった脊髄部位A(青),B(水色)及びAとBを同時刺激した際の力ベクトル(赤)。AとBの線形和で想定されるベクトル(オレンジ)。同時刺激による力ベクトルの方向は個々の脊髄部位の線形和で表現できるが、その大きさは線形和よりはるかに大きい。従って、「運動モジュール」は運動の大きさと方向を別々に制御していることが分かった。
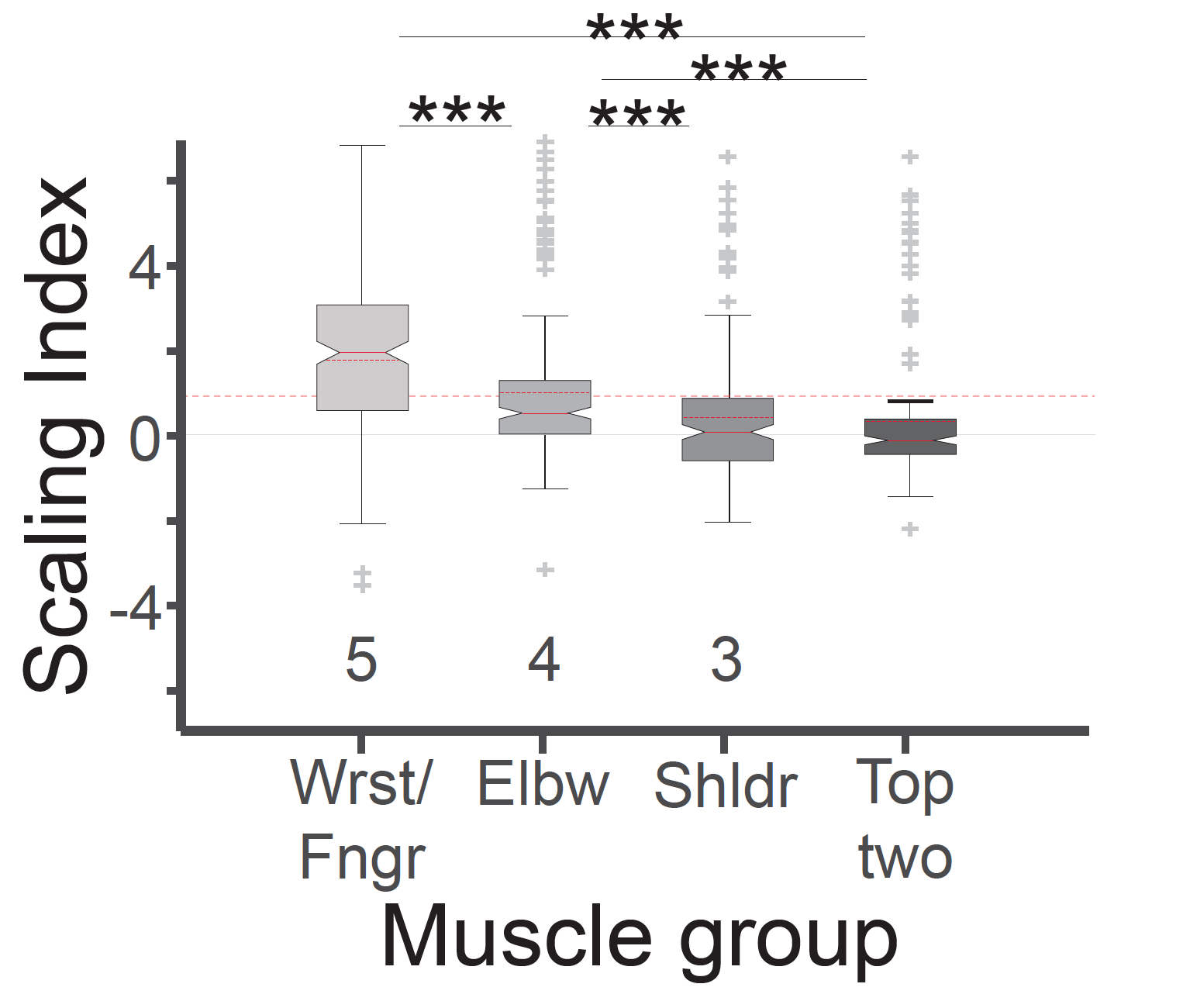
図7:2点同時刺激の超線形和現象の筋肉による相違。超線形和の程度を示す指標(Scaling Index)は手首や指の筋肉において最も大きかった。
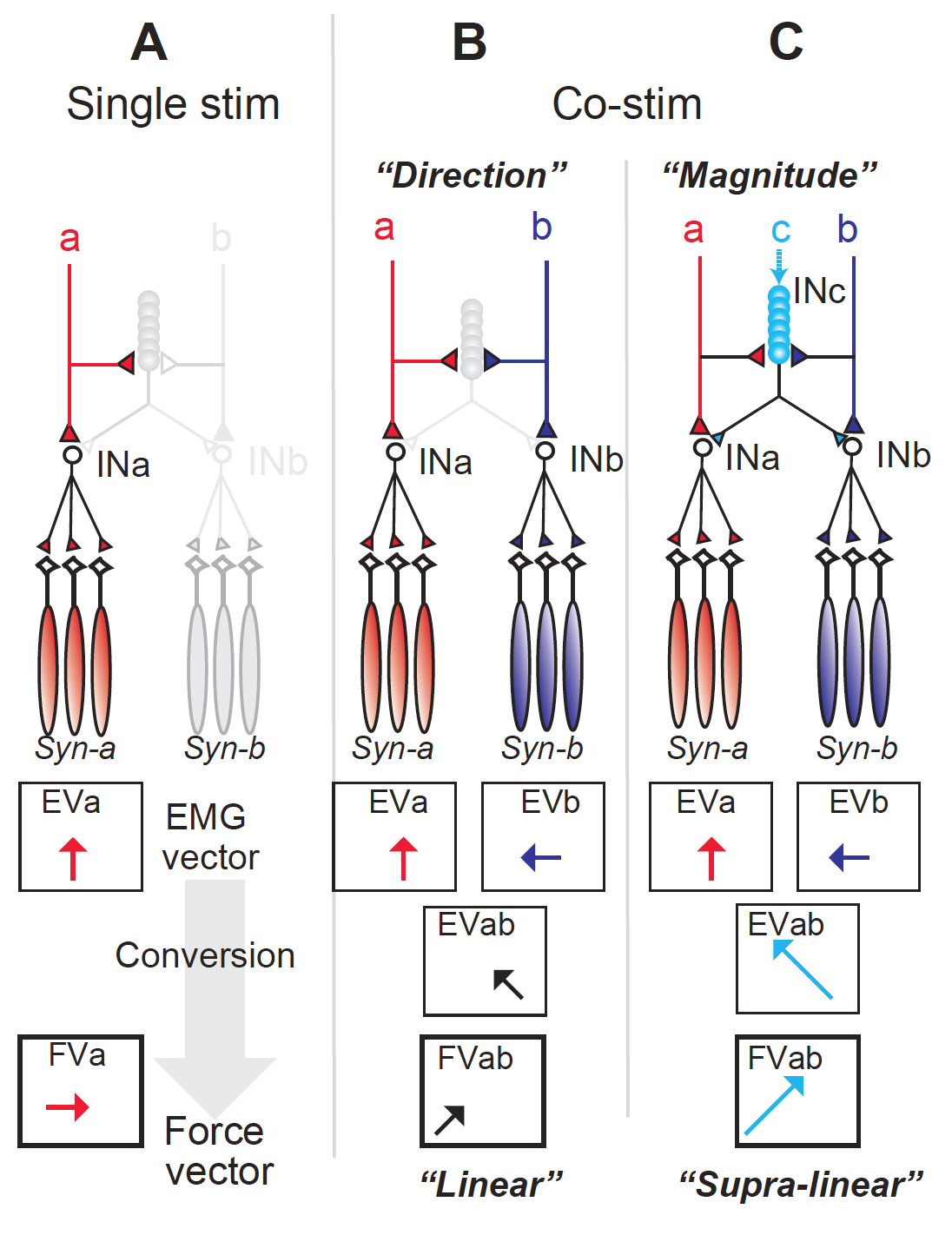
以上の結果から、霊長類の腕の運動の方向と大きさは、脊髄に存在する同じ運動モジュールを異なった様式で利用することによって効率的に制御されていることが示唆されました(図8)。まず運動の方向の制御は、脳からの指令が異なった運動方向を生み出す二つの筋肉のセットを活動させる運動モジュール(脊髄介在ニューロン群(INa、INb))を適切な割合で刺激することによって実現します。すなわち、INa、 INbからの運動出力の線形和*4が、実行したい運動方向と一致するようにそれぞれを活動させるという方法です。この戦略は、カエルや齧歯類でも認められていることから、脊椎動物共通の運動制御戦略と考えることができます。
一方、私たちの日常生活では、同じ方向の運動であっても異なる大きさの力を出す必要がある場合があります。例えば、水がいっぱいに満たされているグラスと、少ししか入っていないグラスを持ち上げて口に運ぶ場合、グラスをテーブルから口まで運ぶために必要な軌道(運動方向)は同一ですが、持ち上げるために必要な筋力(運動の大きさ)は異なります。上記のシステムを用いれば、図中a bを活動させることで運動の方向を決めておきながら、水の量に合わせてまた別の上位モジュールcを活動させることにより、運動の方向と大きさを別々に制御できることになります。
齧歯類やカエルに比較して、霊長類は多様な外部環境を腕を用いて操作することが日常生活において求められています。本研究で新たに発見された運動モジュールと、それを用いた運動方向とサイズの独立制御は、このような霊長類に特有の多機能な手指や手首の運動を効率的に行うために有効に使われていると考えられます。
今後の展望
本研究により、霊長類の腕運動が脊髄に存在する運動モジュールによって効率的に制御されている可能性が、初めて実験的に証明されました。この研究結果によって、今後、ヒトの四肢運動を運動モジュール仮説を土台とした解析や解釈が一層進むと想定されます。工学の分野では、より複雑なロボットの動き制御が運動モジュールを用いた制御によって実現すると期待されます。一方、臨床医学の分野では神経変性疾患や脳卒中などで引き起こされる運動障害を、運動モジュール仮説によって解析することにより、新たな診断方法や治療方法が生み出されることが期待されます。
用語解説
*1運動モジュール仮説:
身体の運動は膨大な数の筋肉や関節が協調して働くことにより実現するが、これがどのような神経の働きで達成されているかはまだ不明である。この命題に対して、1960年代にロシアの科学者ベルンシュタインが提唱した仮説。「脳は、個々の筋肉や関節をそれぞれ別々に制御しているのではなく、神経系内に想定される運動モジュールを制御している。運動モジュールはいくつかの典型的な協調パタンを表現しており、脳はその組み合わせパタンを調節して運動を作り出している」という考え方。長年この運動モジュールの神経実体は未知であったが、1990年代にアメリカMITのビッツィーたちのグループによって、カエルの脊髄に存在することが証明され、カエルの下肢の動きは基本的に運動モジュールの線形和で説明できることが示されていた。
*2脊髄:
脊椎の内側にあり、脳の信号を身体に運ぶ下行路、四肢などからの感覚を運ぶ一次感覚神経、筋肉への指令経路である運動神経が通る白質と、それらの情報を統合する灰白質から構成される。灰白質には反射、歩行、姿勢維持などを無意識下で行うための神経回路が存在することが知られている。脊髄介在ニューロンはその神経回路の中心的実体である。
*3電気刺激による運動の誘発:
脊髄灰白質には、筋線維に直接投射して筋活動を生成する運動ニューロン細胞が存在する。またその運動ニューロン細胞に対して、感覚や脳からの指令を統合した情報を送る介在ニューロンが存在する。脊髄灰白質に電極を入れて、微弱電流で刺激を行うと、これらの介在ニューロンや運動ニューロンが興奮して筋活動や力を作り出す。したがって、誘発された力や筋活動は、脊髄介在ニューロンにおける感覚や運動の統合の特徴を反映している。
*4力ベクトルの線形和と超線形和:
「運動モジュール」を構成する神経細胞群はそれぞれ固有のパタンで筋肉を興奮させ、それによって固有の力ベクトルを発生する。もし、脳が二つのモジュールを組み合わせて使っているなら、二つの「運動モジュール」(二種類の神経細胞群)を電気刺激で同時に興奮させた場合、それぞれの神経細胞群に特有の力ベクトルの単純な足し算(線形和)で推定される力ベクトルが誘発されるはずである。実験結果から、力ベクトルの方向については予想通りの線形和となった。一方、力ベクトルの大きさは、線形和で予想させる大きさの数倍から数十倍の大きさであった。このベクトル線形和で予想されるベクトルサイズよりもはるかに大きな力ベクトルを超線形和と呼んだ。
原論文情報
・論文名:Forelimb force direction and magnitude independently controlled by spinal modules in the macaque“
・著者:Amit Yaron, David Kowalski, 矢口 博彬、武井智彦、関和彦
・掲載誌:Proceedings of the National Academy of Sciences of the United States of America
・DOI:10.1073/pnas.1919253117
・URL:https://doi.org/10.1073/pnas.1919253117
助成金
本成果は、主に以下の研究助成を受けて行われました。
• 文部科学省科学研究費助成金 18020030, 18047027, 26120003, 19H05724, 19H01092,
• 科学技術振興機構「さきがけ」
• 米国NSF grant OISE-1108063.
お問い合わせ先
【研究に関するお問い合わせ】
国立研究開発法人 国立精神・神経医療研究センター
神経研究所 モデル動物開発研究部
関和彦(せき かずひこ)
TEL: 042-346-1724
FAX: 042-346-1754
E-mail: seki(a)ncnp.go.jp
【報道に関するお問い合わせ】
国立研究開発法人国立精神・神経医療研究センター
総務課 広報係
〒187-8551 東京都小平市小川東町4-1-1
TEL: 042-341-2711(代表) FAX: 042-344-6745
国立大学法人京都大学
総務部広報課 国際広報室
〒606-8501 京都市左京区吉田本町
TEL:075-753-5729 FAX:075-753-2094
※E-mailは上記アドレス(a)の部分を@に変えてご使用ください



{kind=link}